Отличие эукариотов от прокариотов хлоропласты. Сравнение прокариот и эукариот. Сравнительная характеристика клеток растений, животных, бактерий, грибов
ОСНОВЫ БОТАНИКИ. НИЗШИЕ РАСТЕНИЯ
Строение эукариотической клетки. Отличие эукариот от
прокариот. Особенности строения и функционирования
растительной клетки. Фотосинтез. Систематика и общая
характеристика водорослей. Водоросли одноклеточные и
многоклеточные, их роль в природных процессах.
Строение эукариотической клетки. Отличия эукариот от прокариот
Эукарио́ты (лат. Eukaryota от др.-греч. εὖ- «хорошо» и κάρυον «ядро»),или я́дерные, - домен (надцарство) живых организмов, клетки которых
содержат ядро. Животные, растения, грибы, а также группы организмов под общим
названием протисты - все являются эукариотическими организмами.
Типичной эукариотической клетки не существует,
но можно выделить общий план строения.
Оболочку, покрывающую клетку снаружи,
называют клеточной мембраной. Внутри клетки
часто встречаются пузырьки, оболочка которых
очень похожа на клеточную мембрану. Их
называют мембранными пузырьками,
или вакуолями.
Различные части клетки называются органоидами: ядра, эндоплазматическая сеть
(ЭПС), комплекс Гольджи, митохондрии, центриоли (вместе они имеют название
"клеточный центр"). Внутреннее содержимое клетки, за исключением ядра,
называют цитоплазмой.Схема типичной клетки животного. 1. Ядрышко 2. Ядро 3. Рибосома 4.Везикула 5.
Шероховатый (гранулярный) эндоплазматический ретикулум 6. Аппарат
Гольджи 7.Клеточная стенка 8. Гладкий (агранулярный) эндоплазматический
ретикулум 9. Митохондрия 10. Вакуоль11. Гиалоплазма 12. Лизосома
13. Центросома (Центриоль)
Согласно наиболее распространённым гипотезам, эукариоты появились 1,5-2 млрд лет
назад. Важную роль в эволюции эукариот сыграл симбиогенез - симбиоз между
эукариотической клеткой, видимо, уже имевшей ядро и способной к фагоцитозу, и
поглощёнными этой клеткой бактериями - предшественниками митохондрий и пластид.
Отличия эукариот от прокариот
1. Расположением генетического аппарата в клетке. Генетический аппарат всехэукариот находится в ядре и защищён ядерной оболочкой. ДНК эукариот
линейная, а у прокариот ДНК кольцевая и находится в особой области клетки -
нуклеоиде, который не отделён мембраной от остальной цитоплазмы. ДНК связана с
белками-гистонами и другими белками хромосом, которых нет у бактерий.
2. В жизненном цикле эукариот обычно присутствуют две ядерные фазы
(гаплофаза и диплофаза). Первая фаза характеризуется гаплоидным (одинарным)
набором хромосом, далее, сливаясь, две гаплоидные клетки (или два ядра)
образуют диплоидную клетку (ядро), содержащую двойной (диплоидный) набор
хромосом. Иногда при следующем делении, а чаще спустя несколько делений
клетка вновь становится гаплоидной. Такой жизненный цикл и в целом
диплоидность для прокариот не характерны.3. Наличие у эукариотических клеток особых органелл, имеющих свой
генетический аппарат, размножающихся делением и окружённых мембраной. Эти
органеллы - митохондрии и пластиды.
Прокариоты характеризуются малым количеством органелл, и ни одна из них не
окружена двойной мембраной. В клетках прокариот нет эндоплазматического
ретикулума, аппарата Гольджи, лизосом.
4. Наличие у эукариот эндоцитоза, в том числе у многих групп - фагоцитоза.
Этот процесс обеспечивает в организме важную защитную функцию. Появление
фагоцитоза у эукариот скорее всего связано с размерами. Размеры
прокариотических клеток несоизмеримо меньше, и поэтому в процессе
эволюционного развития эукариот у них возникла проблема снабжения организма
большим количеством пищи. Как следствие среди эукариот появляются первые
настоящие, подвижные хищники.
5. Различия в организации цитоскелета. Некоторое время считалось, что цитоскелет свойственен только эукариотам, но в середине 1990-х гг. белк
5. Различия в организации цитоскелета. Некоторое время считалось, чтоцитоскелет свойственен только эукариотам, но в середине 1990-х гг.
белки, гомологичные основным белкам цитоскелета эукариот, были обнаружены и у
бактерий.
Именно наличие специфическим образом устроенного цитоскелета позволяет
эукариотам создать систему подвижных внутренних мембранных органоидов. Кроме
того, цитоскелет позволяет осуществлять эндо- и экзоцитоз (как предполагается,
именно благодаря эндоцитозу в эукариотных клетках появились внутриклеточные
симбионты, в том числе митохондрии и пластиды).
Другая важнейшая функция цитоскелета эукариот - обеспечение деления ядра
(митоз и мейоз) и тела (цитотомия) эукариотной клетки (деление прокариотических
клеток организовано проще).
Различия в строении цитоскелета объясняют и другие отличия про- и эукариот -
например, постоянство и простоту форм прокариотических клеток и значительное
разнообразие формы и способность к её изменению у эукариотических, а также
относительно большие размеры последних.Особенности строения и функционирования растительной клетки
Растения – это фотоавтотрофные эукариоты.
Это значит, что растения создают органические
вещества из неорганических соединений –
диоксида углерода (СО2) и воды (Н2О) под
действием света.
Царство Растения включает в себя около 400 000
видов и делится на подцарства – низшие
растения (водоросли) и высшие растения –
споровые (мхи, плауны, хвощи, папоротники) и
семенные (голосемянные и покрытосемянные).
Водоросли – это растения, у которых
тело представлено одной клеткой или
нерасчлененным на органы слоевищем,
т. е. у водорослей клетки не
дифференцированы.
Тело высших растений в той или иной
мере расчленено на органы – корень,
стебель, лист.
1. Клетки растений имеют плотные целлюлозные стенки, основным компонентом которой является целлюлоза. 2. В клеточных стенках растений сущ
Особенности строения растительной клеткиРастительная клетка
1. Клетки растений имеют плотные
целлюлозные стенки, основным компонентом которой
является целлюлоза.
2. В клеточных стенках растений существуют углубления -
поры, через которые проходят цитоплазматические
канальца - плазмодесмы, осуществляющие контакт
соседних клеток и обмен веществами между ними.
3. В клетках находятся зелёные пластиды -
хлоропласты, в них зелёный пигмент хлорофилл
(обеспечивает фотосинтез).
4. В основном ведут прикреплённый образ жизни.
5. Запасные вещества в клетках накапливаются в
виде крахмала.
Растут в течение всей жизни.
Жизнедеятельность регулируется фитогормонами.
Аристотель поместил растения в
промежуточное состояние между
неодушевлёнными предметами и
животными. Он определил растения как
живые организмы, которые не способны
самостоятельно передвигаться (в
противоположность животным).
Хлоропласты в
клетках листаСтроение клетки растений
Фотосинтез
Фотосинтез - синтез органических веществ из углекислого газа и воды собязательным использованием энергии света:
6СО2 + 6Н2О + Qсвета → С6Н12О6 + 6О2.
Общая характеристика процесса фотосинтеза
Процесс фотосинтеза состоит из двух фаз и нескольких
этапов, которые идут последовательно.
I Световая фаза
1. Фотофизический этап – происходит во внутренней
мембране хлоропластов и связан с поглощением
солнечной энергии пигментными системами.
2. Фотохимический этап – проходит во внутренней
мембране хлоропластов и связан с преобразованием
солнечной энергии в химическую энергию АТФ и
НАДФН2 и фотолизом воды.
II Темновая фаза
3. Биохимический этап или цикл Кальвина – проходит в
строме хлоропластов. На этом этапе углекислый газ
восстанавливается до углеводов.
В ходе эволюции растений сформировался специализированный
фотосинтеза – лист, органоиды фотосинтеза - хлоропласты
орган
Пласти́ды (от др.-греч. πλαστός - вылепленный) - полуавтономные органеллы высших растений,
водорослей и некоторых фотосинтезирующих простейших. Пластиды имеют от двух до четырёх
мембран, собственный геном и белоксинтезирующий аппарат.Ультраструктура хлоропласта:
1. внешняя мембрана
2. межмембранное пространство
3. внутренняя мембрана (1 + 2 + 3: оболочка)
4. строма (жидкость)
5. тилакоид с просветом (люменом) внутри
6. мембрана тилакоида
7. грана (стопка тилакоидов)
8. тилакоид (ламелла)
9. зерно крахмала
10. рибосома
11. пластидная ДНК
12. пластоглобула (капля жира)
Функционально фотосинтез приурочен к специализированным органеллам –
хлоропластам. Хлоропласты высших растений имеют форму двояко выпуклой
линзы, которая наиболее удобна для поглощения солнечных лучей. Внутри
хлоропластов находится однородное вещество – строма, пронизанная системой
параллельно расположенных мембран, имеющих вид плоских мешочков тилакоидов, они собраны в стопки, называемые гранами. В тилакоидах гран
находятся хлорофилл и каротиноиды, в строме – молекулы ДНК, рибосомы, капли
липидов, крахмальные зёрна и др. включения.
В мембраны тилакоидов хлоропластов встроены фотосинтетические пигменты: хлорофиллы и каротиноиды. Существует несколько разных типов хло
В мембраны тилакоидов хлоропластов встроены фотосинтетические пигменты:хлорофиллы и каротиноиды. Существует несколько разных типов хлорофилла (a, b,
c, d), хлоропласты высших растений содержат хлорофилл а и хлорофилл b.
В молекуле хлорофилла можно выделить порфириновую «головку» с атомом
магния в центре и фитольный «хвост». Порфириновая «головка» представляет собой
плоскую структуру, является гидрофильной и поэтому лежит на той поверхности
мембраны, которая обращена к водной среде стромы. Фитольный «хвост» -
гидрофобный и за счет этого удерживает молекулу хлорофилла в мембране.
Хлорофиллы поглощают красный и сине-фиолетовый
свет, отражают зеленый и поэтому придают
растениям характерную зеленую окраску.
Молекулы хлорофилла в мембранах тилакоидов
организованы в фотосистемы.
У растений и синезеленых водорослей имеются
фотосистема-1 и фотосистема-2, у фотосинтезирующих
бактерий - фотосистема-1. Только фотосистема-2
может разлагать воду с выделением кислорода и
отбирать электроны у водорода воды.Каротиноиды - это желтые, оранжевые, красные или коричневые пигменты, сильно
поглощающие свет в сине-фиолетовой области. Они называются вспомогательными
пигментами, поскольку поглощенную ими световую энергию они переносят на
хлорофилл. В спектре поглощения каротиноидов обнаруживаются три пика в синефиолетовой области. Помимо своей функции как вспомогательных пигментов
каротиноиды защищают хлорофиллы от избытка света и от окисления
кислородом, образующимся в процессе фотосинтеза. Они хорошо замаскированы
зелеными хлорофиллами, но становятся видны в листьях до начала листопада,
поскольку хлорофиллы разрушаются первыми.1. Фотофизический этап
Световая фаза происходит в мембранах тилакоидов только в присутствии света при
участии хлорофилла, белков-переносчиков электронов и фермента - АТФ-синтетазы.
Под действием кванта света электроны хлорофилла возбуждаются, покидают
молекулу и попадают на внешнюю сторону мембраны тилакоида, которая в итоге
заряжается отрицательно. Окисленные молекулы хлорофилла восстанавливаются,
отбирая электроны у воды, находящейся во внутритилакоидном пространстве. Это
приводит к распаду или фотолизу воды:
Н2О + Qсвета → Н+ + ОН-.
Ионы гидроксила отдают свои электроны, превращаясь в
реакционноспособные радикалы ОН:
ОН- → ОН + е-.
Радикалы ОН объединяются, образуя воду и свободный кислород:
4НО → 2Н2О + О2.
Кислород при этом удаляется во внешнюю среду, а протоны накапливаются внутри
тилакоида в «протонном резервуаре». В результате мембрана тилакоида с одной
стороны за счет Н+ заряжается положительно, с другой за счет электронов -
отрицательно.
2. Фотохимический этап
Когда разность потенциалов между наружной и внутренней сторонами мембраны
тилакоида достигает 200 мВ, протоны проталкиваются через каналы АТФсинтетазы и происходит фосфорилирование АДФ до АТФ; атомарный водород
идет на восстановление специфического переносчика
НАДФ+ (никотинамидадениндинуклеотидфосфат) до НАДФ·Н2:
2Н+ + 2е- + НАДФ → НАДФ·Н2.
Таким образом, в световую фазу происходит фотолиз воды, который сопровождается
тремя важнейшими процессами: 1) синтезом АТФ; 2) образованием НАДФ·Н2; 3)
образованием кислорода.
Кислород диффундирует в атмосферу, АТФ и НАДФ·Н2транспортируются в строму
хлоропласта и участвуют в процессах темновой фазы
1 - строма хлоропласта; 2 - тилакоид граны.
Эта фаза протекает в строме хлоропласта. Реакции темновой фазы представляют собой цепочку последовательных преобразований углекислого г
Темновая фазаЭта фаза протекает в строме хлоропласта. Реакции темновой фазы представляют
собой цепочку последовательных преобразований углекислого газа (поступает из
воздуха), приводящую к образованию глюкозы и других органических веществ.
Первая реакция в этой цепочке - фиксация углекислого газа; акцептором
углекислого газа является пятиуглеродный сахар рибулозобифосфат (РиБФ);
катализирует реакцию фермент рибулозобифосфат-карбоксилаза (РиБФкарбоксилаза). В результате карбоксилирования рибулозобисфосфата образуется
неустойчивое шестиуглеродное соединение, которое сразу же распадается на две
молекулы фосфоглицериновой кислоты (ФГК).
Затем происходит цикл реакций, в которых
через ряд промежуточных продуктов
фосфоглицериновая кислота преобразуется в
глюкозу. В этих реакциях используются энергии
АТФ и НАДФ·Н2, образованных в световую
фазу; цикл этих реакций получил название
«цикл Кальвина»:
6СО2 + 24Н+ + АТФ → С6Н12О6 + 6Н2О.
Кроме глюкозы, в процессе фотосинтеза
образуются другие мономеры сложных
органических соединений - аминокислоты,
глицерин и жирные кислоты, нуклеотиды.Систематика и общая характеристика водорослей
Во́доросли (лат. Algae) - гетерогенная экологическая группа преимущественно
фотоавтотрофных одноклеточных, колониальных или многоклеточных
организмов, обитающих, как правило, в водной среде, в систематическом отношении
представляющая собой совокупность многих отделов.
Вступая в симбиоз с грибами, эти организмы в ходе эволюции образовали совершенно
новые организмы -лишайники.
На основании различий в наборе пигментов, структуре хроматофора, особенностей морфологии
и биохимии (состав клеточных оболочек, типы запасных питательных веществ) большинством
отечественных систематиков выделяется 11 отделов водорослей
Прокариоты, или Доядерные
Царство Бактерии
Подцарство Цианобактерии
Отдел Сине-зелёные водоросли
Эукариоты, или Ядерные
Царство Растения
Подцарство Водоросли
Отдел Зелёные водоросли
Отдел Эвгленовые водоросли
Отдел Золотистые водоросли
Отдел Жёлто-зелёные
водоросли
Отдел Диатомовые водоросли
Отдел Динофитовые водоросли
Отдел Криптофитовые
водоросли
Отдел Бурые водоросли
Подцарство Багрянки
Отдел Красные водорослиНекоторые водоросли способны к гетеротрофии, как осмотрофной (поверхностью
клетки), например жгутиконосцы, так и путём заглатывания через клеточный рот
(эвгленовые, динофитовые).
Размеры водорослей колеблются от долей микрона (некоторые диатомеи) до 30-50м
(бурые водоросли - ламинария, саргассум).
Среди одноклеточных есть колониальные формы, когда отдельные клетки тесно
связаны между собой (соединены через плазмодесмы или погружены в общую слизь).
Среда их обитания преимущественно водная, но встречаются они в почве, на
коре деревьев и даже в снегу – снежная хламидомонада. Скопления этой
водоросли придают таящему снегу различные оттенки – от красного до зеленого.
эвглена
Одноклеточные водоросли: 1 – хламидомонада;
2 – хлорелла; 3 – нитчатая водоросль спирогира;
4 – колониальная водоросль вольвокс;
многоклеточные водоросли: 5 – ламинария; 6 – порфира
хламидомонадаВодоросли являются наиболее древней группой растений. Они прошли длительный
эволюционный путь, приспосабливаясь к различным сменявшимся условиям на Земле.
Тело водорослей называется талломом, или слоевищем. У некоторых водорослей есть
ризоиды - нитевидные выросты, в основном предназначенные для прикрепления к
субстрату. Могут выполнять функцию всасывания воды и минеральных веществ.
Обитая в водной среде, они поглощают питательные вещества всей поверхностью.
Вода поглощает и рассеивает свет, поэтому по мере погружения освещенность падает.
Волны красной части спектра практически не проникают на глубину свыше 12 м. А
именно в этой области спектра "работает" хлорофилл. Поэтому для лучшего
обеспечения фотосинтеза у многих групп водорослей появились дополнительные
пигменты, поглощающие свет в синей области спектра. Для каждого отдела
водорослей характерен свой набор пигментов, что отражается в их названиях.Водоросли одноклеточные и многоклеточные,
роль в природных процессах
Диатомовые водоросли. Их тело состоит из одной клетки (встречаются колонии),
которая заключена в прозрачный панцирь. Причем панцири бывают украшены
различными рисунками, и по этим рисункам водоросли и различаются. Панцирь
состоит из кремнезема, они прозрачны для света, поэтому водоросли могут
осуществлять фотосинтез. Когда водоросли отмирают, панцири осаждаются на дно
водоема и образуют породу диатомит.
Одноклеточные водоросли: диатомовые и эвгленовые
Эвгленовые водоросли: ботаники относят их к растениям, а зоологи – к простейшим
животным (содержат хлорофилл – признак растений; могут двигаться – животные).
Кроме того, они могут терять хлорофилл и питаться уже готовыми органическими
веществами (гетеротрофы).
Зеленые водоросли не имеют дополнительных пигментов, поэтому их окраску определяет хлорофилл. Именно эта группа водорослей дала начало вы
Зеленые водоросли не имеют дополнительных пигментов, поэтому их окраскуопределяет хлорофилл. Именно эта группа водорослей дала начало высшим
растениям. Они широко распространены в пресных и морских водах, встречаются
также на суше в увлажненных местах: в почве, на коре деревьев, на камнях. Размеры
их варьируют от нескольких микрометров до метров.
Они представлены различными жизненными формами: одноклеточными,
колониальными, нитчатыми и многоклеточными.
Представителями одноклеточных водорослей являются хламидомонада и
хлорелла.
СТРОЕНИЕ ХЛАМИДОМОНАДЫ
Хламидомонада - округлая клетка, вытянутая с
переднего конца. На нем находится пара жгутиков,
за счет которых она передвигается. Снаружи клетка
покрыта клеточной стенкой. В центре клетки
находится гаплоидное ядро. Единственная крупная
пластида, называемая хроматофор, располагается
по периферии клетки, делая ее окрашенной. В клетке
имеется обычный набор органелл. Кроме того, на
переднем конце располагается пара
сократительных вакуолей, выводящих из клетки
избыток воды.
В условиях неравномерного освещения хламидомонада всегда плывет на свет. Это
явление называется положительным фототаксисом. Для его осуществления у
хламидомонады есть специальный органоид, видимый как маленькая красная точка в
основании жгутиков. Он называется стигма, или глазок.
Размножение и жизненный цикл хламидомонады. Жизненный цикл хламидомонады идет с чередованием гаплоидной и диплоидной форм. В благоприятн
Размножение и жизненный цикл хламидомонады. Жизненный циклхламидомонады идет с чередованием гаплоидной и диплоидной форм. В
благоприятных условиях хламидомонада быстро размножается бесполым путем.
Достигнув определенных размеров, клетка отбрасывает жгуты и округляется.
Происходит, в зависимости от вида, 1, 2 или 3 митотических деления ядра. Под
оболочкой материнской клетки образуется 2, 4 или 8 мелких клеток, имеющих пару
жгутиков. Оболочка материнской клетки разрывается, и мелкие клетки, называемые
зооспорами, выходят в среду. Они растут и превращаются во взрослых
хламидомонад.
В неблагоприятных условиях у
хламидомонады начинается половой
процесс. Внутри родительских клеток
формируются подвижные гаметы, которые
выходят в воду. Гаметы, происходящие из
разных родительских клеток, соединяются
попарно и образуют зиготу. Она
покрывается плотной оболочкой и
превращается в зигоцисту, способную
переживать неблагоприятные условия. При
наступлении благоприятных условий в
зигоцисте происходит мейоз, и из нее
выходят 4 зооспоры, вырастающие во
взрослую хламидомонаду
Хлорелла. В отличие от хламидомонады, хлорелла не имеет жгутиков и удерживается в верхних слоях воды за счет низкой плотности. Выглядит она
какзеленая муть в воде - вода «цветет».
пиреноид - особое включение внутри
хлоропласта. Функция - запасание фермента
рибулозобисфосфаткарбоксилаза (рубиско)
и/или центр синтеза сахаров при помощи
этого фермента.
Размножается она только бесполым путем, а неблагоприятные условия переживает
в форме цисты, в которые превращаются обычные клетки. Для хлореллы характерна
высокая скорость фотосинтеза, она богата белками и липидами, благодаря чему ее
выращивают на корм скоту и применяют для регенерации кислорода в космических
аппаратах
Многоклеточные зеленые водоросли самой разнообразной формы: кустистые, нитчатые, слоевидные. Но у всех клетки не дифференцированы.
Ульва, или морской салат – род зелёныхводорослей, включающему около 20 видов. Ульва
растет чаще всего на мелководье, это
светолюбивое и теплолюбивое растение.
Строение. Многоклеточное ярко-зеленое слоевище этой водоросли пластинчатое,
может быть цельное, рассеченное либо ветвистое. Длина таллома составляет, в
среднем, 0,3-1,5 метра. В основании слоевища имеются крупные клетки с ризоидами,
образующие подошву, посредством которой водоросль прикрепляется к подводному
субстрату.
Размножение. Возможен вегетативный и половой способы размножения. Из
фрагментов организма появляется новое растение. От основания ульвы, точнее из ее
подошвы могут развиваться молодые растения.
При половом процессе происходит слияние гамет из
разных талломов (с помощью подвижных клеток, которые
образуются на месте материнского таллома). Жизненный
цикл ульвы включает два поколения - гаметофит и
спорофит, которые выглядят абсолютно одинаково и
имеют сходное строение. Половое размножение может
протекать как изогамия или гетерогамия.
Представителями нитчатых зеленых водорослей являются улотрикс и спирогира. Улотрикс растет в прикрепленном состоянии. Нижняя клетка нити
Представителями нитчатых зеленых водорослей являютсяулотрикс и спирогира.
Улотрикс растет в прикрепленном состоянии. Нижняя клетка нити,
называемая прикрепительной (ризоидальной) клеткой, плотно
врастает в поверхность какого-либо подводного предмета, образует
толстую клеточную стенку, ее цитоплазма отмирает. Остальные
клетки имеют одинаковое строение и способны к делению и
фотосинтезу. За счет их деления водоросль растет в длину.
Улотрикс размножается половым и бесполым путем.
Бесполое размножение улотрикса осуществляется с помощью подвижных 4жгутиковых зооспор. Они образуются путем митотического деления из клеток
средней части нити. Прикрепившись к какой-нибудь поверхности, они сбрасывают
жгуты и делятся митозом в плоскости, параллельной поверхности. Нижняя клетка
превращается в прикрепительную, а верхняя продолжает делиться, образуя нить.
Нити улотрикса могут размножаться фрагментацией.
В неблагоприятных условиях улотрикс
размножается половым путем. В клетках нити
формируются подвижные гаметы. Они,
соединяясь попарно, образуют зиготу, которая
превращается в зигоцисту, переживающую
неблагоприятные условия. В благоприятных
условиях в ней происходит мейоз, и
образовавшиеся гаплоидные клетки дают
начало новым нитям улотрикса.
Спирогира представляет собой длинные плавающие в толще воды нити, состоящие из крупных клеток. Центр клетки занимает крупная центральная
Спирогира представляет собой длинные плавающие в толще воды нити,состоящие из крупных клеток. Центр клетки занимает крупная центральная
вакуоль, цитоплазма находится в пристенном слое и пронизывает вакуоль
отдельными тяжами. Особенность спирогиры: один или несколько
лентовидных хроматофоров, закрученных в спираль, и гаплоидное ядро.
Нить растет за счет деления всех клеток.
При фрагментации нити каждый ее кусочек может
дать начало новой нити - вегетативное
размножение спирогиры. Часто в водоемах
спирогира образует густые сплетения, похожие на
зеленую вату.
Половой процесс - конъюгация - у спирогиры
происходит между обычными клетками двух разных
нитей.
При сближении нитей между ними образуется конъюгационная
трубка. Содержимое одной клетки, принадлежащей к «+»-нити,
перетекает в другую, принадлежащую «–»-нити.
Происходит слияние клеток, а затем и ядер. Формируется
диплоидная зигота, которая окружается плотной оболочкой -
образуется зигоспора.
Зигота делится мейозом, образуя 4 гаплоидные клетки. В дальнейшем 3 из 4 клеток
погибают. Оставшаяся прорастает в гаплоидную нить спирогиры.
Бурые водоросли – это многоклеточные водоросли, обитающие исключительно в соленых водах. К бурым водорослям относится морская капуста ла
Бурые водоросли – это многоклеточные водоросли, обитающие исключительно всоленых водах. К бурым водорослям относится морская капуста ламинария, фукус,
цистозира, саргассум. Они замечательны тем, что накапливают много йода,
содержащегося в морской воде.
Они содержат хлорофилл и, кроме того, желтые и оранжевые пигменты, которые
помогают улавливать свет и осуществлять фотосинтез.
Фукус
Падина павлинья
Ламинария
Цистозира бородатая
С помощью дополнительных пигментов они могут осуществлять фотосинтез на глубине до 30 метров. Они представляют собой крупные растения (до 30
метров вдлину), состоящие из диплоидных клеток. Таллом образует ризоиды для
прикрепления к субстрату. Многие из них растут в приливно-отливной зоне
(литорали) и во время отлива оказываются на суше. Для защиты от высыхания
бурые водоросли образуют много слизистых веществ. Таллом фукуса содержит
многочисленные пузырьки воздуха для увеличения плавучести
В жизненном цикле бурых водорослей наблюдается чередования гаплоидного гаметофита и диплоидного спорофита с преобладанием спорофита. Ра
В жизненном цикле бурых водорослей наблюдается чередования гаплоидногогаметофита и диплоидного спорофита с преобладанием спорофита.
Размножаются бурые водоросли половым и бесполым путем.
Диплоидные растения посредством мейоза образуют гаплоидные клетки. У
одних (род фукус) они становятся гаметами, при слиянии которых образуется
зигота, дающая начало новому растению. У большинства же продуктами мейоза
являются споры, которые дают начало гаплоидной стадии.
Гаплоидная стадия представляет
собой мелкие нитевидные
образования, которые недолго
живут на дне моря. Они
раздельнополы. На них
формируются многоклеточные
половые органы, в которых
образуются гаметы: яйцеклетки и
сперматозоиды. Они, сливаясь,
образуют зиготу, из которой
вырастают крупные диплоидные
растения.
Жизненный цикл ламинарии
Сифоновые водоросли - одни из самых древних групп зеленых водорослей. У них таллом образован, как правило, одной гигантской клеткой. В цито
Сифоновые водоросли - одни из самых древних групп зеленых водорослей. Уних таллом образован, как правило, одной гигантской клеткой. В цитоплазме кроме
одного или нескольких ядер содержится также один или несколько хлоропластов.
Многочисленные хлоропласты обладают дисковидной или веретеновидной
формой; когда хлоропласт один, он имеет сетчатое строение. Примерами таких
водорослей являются каулерпа и ацетабулярия.
каулерпа
ацетабулярия
Ацетабулярия. Нижняя часть одноклеточного слоевища (ризоид) находится в грунте.
В ризоиде расположено ядро. Вверх растет ножка, достигающая в длину
нескольких сантиметров. На ее конце формируется шляпка. Для размножения по
периферии шляпки образуются споры, из которых вырастают новые растения.
Красные водоросли – совершенно особая группа водорослей, которая благодаря особым ферментам способна обитать на больших глубинах (до пят
Красные водоросли – совершенно особая группа водорослей, которая благодаряособым ферментам способна обитать на больших глубинах (до пятисот метров).
Среди багрянок встречаются как одноклеточные, так и
многоклеточные нитчатые и пластинчатые водоросли.
Из 4000 видов только 200 приспособились к жизни в
пресных водоемах и на почве, остальные – обитатели
морей. Окраска красных водорослей определяется
различным количественным содержанием пигментов:
хлорофиллы а и d, каротиноиды и фикобиллины:
красный (фикоэритрин) и синий (фикоцианин).
Причем окраска водорослей различна на разной
глубине, на мелководье они желто-зеленые, затем
розовые и на глубине более 50 м становятся
красными.
Формы таллома багрянок
филлофора ребристая
У берегов Крыма узкой полосой встречается прикрепленная форма
филлофоры ребристой на валунах и скалах на глубинах до 60 м.
Крупный таллом, до 50 см, с плоскими ветвями образует кусты
малиново-красного цвета. Филлофора - важный источник
получения желирующих веществ, в частности агароидов, которые
находят применение в текстильной, пищевой и микробиологической
промышленности.
Харовые водоросли - класс, которые объединяют в себе признаки водорослей и высших растений. По строению они похожи на хвощи. Их тело состо
Харовые водоросли - класс, которые объединяют в себе признаки водорослейи высших растений. По строению они похожи на хвощи. Их тело состоит из
отдельных члеников, ветвеподобных образований. Всего известно не более 700
видов харовых
Высота их таллома составляет обычно 20-30 см, но может
достигать и 1-2 м, боковые ветви ограниченного роста,
расположены мутовками на многоклеточных
узлах. Междоузлия состоят из одной длинной клетки,
которая может обрастать корой из узких клеток.
Хлоропласты зелёные, содержат хлорофиллы a и b, из
дополнительных пигментов - ликопин. Запасное
вещество - крахмал.
Размножение: вегетативное
и половое. Вегетативное
размножение осуществляется
посредством специальных
клубеньков на ризоидах или
звездообразных скоплений клеток на
нижних стеблевых узлах, которые
дают начало новому таллому.
Хара ломкая
Репродуктивные органы харовых водорослей (сверху: оогоний, снизу:антеридий)
Женский половой орган -оогоний и мужской - антеридий многоклеточны и развиваются
у большинства видов на одном растении, но известны и двудомные виды.Пирофитовые водоросли - группа одноклеточных морских (реже пресноводных)
жгутиковых
водорослей,
объединяющая
около
2100
видов
из
двух
подотделов: криптофитовые и динофитовые. Хлоропласты бурые, клетка, как
правило, заключена в панцирь из целлюлозы, нередко причудливой формы.
Большинство пирофитов – автотрофы. Размножаются делением и спорами, половой
процесс наблюдается редко. Пирофитовые водоросли – причина «красных приливов»;
выделяемые множеством этих микроорганизмов токсичные вещества служат причиной
гибели рыб и моллюсков. Другие пирофиты – симбионты радиолярий и коралловых
полипов.
К ним относится ночесветка. Несмотря на микроскопические размеры, её можно
увидеть невооруженным глазом.
Она обладает удивительным свойством - ярко
светиться. Днем это явление незаметно, но ночами
свечение становится очень хорошо видным.
Многие представители пирофитовых водорослей
обладают способностью биолюминесценции. Этот
живой свет имеет множество оттенков, но в
основном бывает серебристо-белым.
Шаровидное тело ночесветки едва достигает 2 мм в диаметре. Имеющиеся жировые
включения обеспечивают «парение» организма в воде. Она лишена оболочки из
клетчатки и не имеет, в отличие от других панцирных жгутиконосцев, хроматофоров, а
потому неспособна к фотосинтезу. Питается ночесветка, заглатывая и
переваривая мелкие виды простейших водорослей, для чего у нее имеется
отверстие, выполняющее функцию рта.Значение водорослей.
1. В основном, все пищевые цепи в морях и океанах начинаются с планктонных
водорослей. Водоросли являются строителями экосистем в шельфовой, прибрежной,
зоне и большую роль некоторые красные водоросли играют в строении коралловых
рифов. Они строятся не только кораллами, но и красными водорослями, в теле
которых тоже может откладываться известь.
2. Поглощение углекислого газа и выделение кислорода.
3. Место обитания рыб.
4. Панцири диатомовых – источник диатомита.
5.Источник агар-агара.
6. Источник йода и минеральных веществ.
7. Удобрение
8. Корм для скота.
По строению клетки живые организмы делят на прокариот и эукариот . Клетки и тех и других окружены плазматической мембраной , снаружи от которой во многих случаях имеется клеточная стенка . Внутри клетки находится полужидкая цитоплазма . Однако клетки прокариот устроены значительно проще, чем клетки эукариот.
Основной генетический материал прокариот (от греч. про – до и карион – ядро) находится в цитоплазме в виде кольцевой молекулы ДНК. Эта молекула (нуклеоид ) не окружена ядерной оболочкой, характерной для эукариот, и прикрепляется к плазматической мембране (рис.1). Таким образом, прокариоты не имеют оформленного ядра. Кроме нуклеоида в прокариотической клетке часто встречается небольшая кольцевая молекула ДНК, называемая плазмидой . Плазмиды могут перемещаться из одной клетки в другую и встраиваться в основную молекулу ДНК.
Некоторые прокариоты имеют выросты плазматической мембраны: мезосомы, ламеллярные тилакоиды, хроматофоры . В них сосредоточены ферменты, участвующие в фотосинтезе и в процессах дыхания. Кроме того, мезосомы ассоциированы с синтезом ДНК и секрецией белка.
Клетки прокариот имеют небольшие размеры, их диаметр составляет 0,3–5 мкм. С наружной стороны плазматической мембраны всех прокариот (за исключением микоплазм) находится клеточная стенка . Она состоит из комплексов белков и олигосахаридов, уложенных слоями, защищает клетку и поддерживает ее форму. От плазматической мембраны она отделена небольшим межмембранным пространством.
В цитоплазме прокариот обнаруживаются только немембранные органоиды рибосомы . По структуре рибосомы прокариот и эукариот сходны, однако рибосомы прокариот имеют меньшие размеры и не прикрепляются к мембране, а располагаются прямо в цитоплазме.

Многие прокариоты подвижны и могут плавать или скользить с помощью жгутиков.
Размножаются прокариоты обычно путем деления надвое (бинарным ). Делению предшествует очень короткая стадия удвоения, или репликации, хромосом. Так что прокариоты – гаплоидные организмы.
К прокариотам относятся бактерии и синезеленые водоросли, или цианобактерии. Прокариоты появились на Земле около 3,5 млрд лет назад и были, вероятно, первой клеточной формой жизни, дав начало современным прокариотам и эукариотам.
Эукариоты (от греч. эу – истинный, карион – ядро) в отличие от прокариот, имеют оформленное ядро, окруженное ядерной оболочкой – двуслойной мембраной. Молекулы ДНК, обнаруживаемые в ядре, незамкнуты (линейные молекулы). Кроме ядра часть генетической информации содержится в ДНК митохондрий и хлоропластов. Эукариоты появились на Земле примерно 1,5 млрд лет назад.
В отличие от прокариот, представленных одиночными организмами и колониальными формами, эукариоты могут быть одноклеточными (например, амеба), колониальными (вольвокс) и многоклеточными организмами. Их делят на три больших царства: Животные, Растения и Грибы.
Диаметр клеток эукариот составляет 5–80 мкм. Как и прокариотические клетки, клетки эукариот окружены плазматической мембраной , состоящей из белков и липидов. Эта мембрана работает как селективный барьер, проницаемый для одних соединений и непроницаемый для других. Снаружи от плазматической мембраны расположена прочная клеточная стенка , которая у растений состоит главным образом из волокон целлюлозы, а у грибов – из хитина. Основная функция клеточной стенки – обеспечение постоянной формы клеток. Поскольку плазматическая мембрана проницаема для воды, а клетки растений и грибов обычно соприкасаются с растворами меньшей ионной силы, чем ионная сила раствора внутри клетки, вода будет поступать внутрь клеток. За счет этого объем клеток будет увеличиваться, плазматическая мембрана начнет растягиваться и может разорваться. Клеточная стенка препятствует увеличению объема и разрушению клетки.
У животных клеточная стенка отсутствует, но наружный слой плазматической мембраны обогащен углеводными компонентами. Этот наружный слой плазматической мембраны клеток животных называют гликокаликсом . Клетки многоклеточных животных не нуждаются в прочной клеточной стенке, поскольку есть другие механизмы, обеспечивающие регуляцию клеточного объема. Так как клетки многоклеточных животных и одноклеточные организмы, живущие в море, находятся в среде, в которой суммарная концентрация ионов близка к внутриклеточной концентрации ионов, клетки не набухают и не лопаются. Одноклеточные животные, живущие в пресной воде (амеба, инфузория туфелька), имеют сократительные вакуоли, которые постоянно выводят наружу поступающую внутрь клетки воду.
Структурные компоненты эукариотической клетки
Внутри клетки под плазматической мембраной находятся цитоплазма . Основное вещество цитоплазмы (гиалоплазма) представляет собой концентрированный раствор неорганических и органических соединений, главными компонентами которого являются белки. Это коллоидная система, которая может переходить из жидкого в гелеобразное состояние и обратно. Значительная часть белков цитоплазмы является ферментами, осуществляющими различные химические реакции. В гиалоплазме располагаются органоиды, выполняющие в клетке различные функции. Органоиды могут быть мембранными (ядро, аппарат Гольджи, эндоплазматический ретикулум, лизосомы, митохондрии, хлоропласты) и немембранными (клеточный центр, рибосомы, цитоскелет).
Мембранные органоидысновным компонентом мембранных органоидов является мембрана . Биологические мембраны построены по общему принципу, но химический состав мембран разных органоидов различен. Все клеточные мембраны – это тонкие пленки (толщиной 7–10 нм), основу которых составляет двойной слой липидов (бислой), расположенных так, что заряженные гидрофильные части молекул соприкасаются со средой, а гидрофобные остатки жирных кислот каждого монослоя направлены внутрь мембраны и соприкасаются друг с другом (рис. 3). В бислой липидов встроены молекулы белков (интегральные белки мембраны) таким образом, что гидрофобные части молекулы белка соприкасаются с жирнокислотными остатками молекул липидов, а гидрофильные части экспонированы в окружающую среду. Кроме этого часть растворимых (немембранных белков) соединяется с мембраной в основном за счет ионных взаимодействий (периферические белки мембраны). Ко многим белкам и липидам в составе мембран присоединены также углеводные фрагменты. Таким образом, биологические мембраны – это липидные пленки, в которые встроены интегральные белки.
Одна из основных функций мембран – создание границы между клеткой и окружающей средой и различными отсеками клетки. Липидный бислой проницаем в основном для жирорастворимых соединений и газов, гидрофильные вещества переносятся через мембраны с помощью специальных механизмов: низкомолекулярные – с помощью разнообразных переносчиков (каналов, насосов и др.), а высокомолекулярные – с помощью процессов экзо- и эндоцитоза (рис. 4).
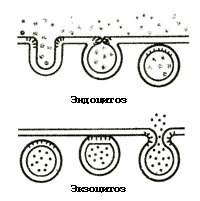
Рис. 4. Схема переноса веществ через мембрану
При эндоцитозе определенные вещества сорбируются на поверхности мембраны (за счет взаимодействия с белками мембраны). В этом месте образуется впячивание мембраны внутрь цитоплазмы. Затем от мембраны отделяется пузырек, внутри которого содержится переносимое соединение. Таким образом, эндоцитоз – это перенос в клетку высокомолекулярных соединений внешней среды, окруженных участком мембраны. Обратный процесс, то есть экзоцитоз – это перенос веществ из клетки наружу. Он происходит путем слияния с плазматической мембраной пузырька, заполненного транспортируемыми высокомолекулярными соединениями. Мембрана пузырька сливается с плазматической мембраной, а его содержимое изливается наружу.
Каналы, насосы и другие переносчики – это молекулы интегральных белков мембраны, обычно образующие в мембране пору.
Кроме функций разделения пространства и обеспечения избирательной проницаемости мембраны способны воспринимать сигналы. Эту функцию осуществляют белки-рецепторы, связывающие сигнальные молекулы. Отдельные белки мембраны являются ферментами, осуществляющими определенные химические реакции.
Ядро – крупный органоид клетки, окруженный ядерной оболочкой и имеющий обычно шаровидную форму. Ядро в клетке одно, и хотя встречаются многоядерные клетки (клетки скелетных мышц, некоторых грибов) или не имеющие ядра (эритроциты и тромбоциты млекопитающих), но эти клетки возникают из одноядерных клеток-предшественников.
Основная функция ядра – хранение, передача и реализация генетической информации . Здесь происходит удвоение молекул ДНК, в результате чего при делении дочерние клетки получают одинаковый генетический материал. В ядре с использованием в качестве матрицы отдельных участков молекул ДНК (генов) происходит синтез молекул РНК: информационных (иРНК), транспортных (тРНК) и рибосомальных (рРНК), необходимых для синтеза белка. В ядре осуществляется сборка субъединиц рибосом из молекул рРНК и белков, которые синтезируются в цитоплазме и переносятся в ядро.
Ядро состоит из ядерной оболочки, хроматина (хромосом), ядрышка и нуклеоплазмы (кариоплазмы).

Рис. 5. Структура хроматина: 1 – нуклеосома, 2 – ДНК
Под микроскопом внутри ядра видны зоны плотного вещества – хроматина. В неделящихся клетках он равномерно заполняет объем ядра или конденсируется в отдельных местах в виде более плотных участков и хорошо окрашивается основными красителями. Хроматин представляет собой комплекс ДНК и белков (рис. 5), большей частью положительно заряженных гистонов .
Количество молекул ДНК в ядре равно числу хромосом. Количество и форма хромосом являются уникальной характеристикой вида. В состав каждой из хромосом входит одна молекула ДНК, состоящая из двух связанных между собой нитей и имеющая вид двойной спирали толщиной 2 нм. Длина ее значительно превышает диаметр клетки: она может достигать нескольких сантиметров. Молекула ДНК заряжена отрицательно, поэтому сворачиваться (конденсироваться) она может только после связывания с положительно заряженными белками-гистонами (рис. 6).
Сначала двойная нить ДНК закручивается вокруг отдельных блоков гистонов, в каждый из которых входит 8 молекул белка, образуя структуру в виде «бусин на нитке» толщиной около 10 нм. Бусины называются нуклеосомами. В результате формирования нуклеосом длина молекулы ДНК уменьшается примерно в 7 раз. Далее нить с нуклеосомами сворачивается, формируя структуру в виде каната толщиной около 30 нм. Затем такой канат, изогнутый в виде петель, прикрепляется к белкам, образующим основу хромосомы. В результате образуется структура с толщиной около 300 нм. Дальнейшая конденсация этой структуры приводит к образованию хромосомы.
В период между делениями хромосома частично разворачивается. В результате этого отдельные участки молекулы ДНК, которые должны экспрессироваться в данной клетке, освобождаются от белков и вытягиваются, что делает возможным считывание с них информации путем синтеза молекул РНК.
Ядрышко – это тип матричной ДНК, отвечающей за синтез рРНК и собранной в отдельных участках ядра. Ядрышко – наиболее плотная структура ядра, оно не является отдельным органоидом, а представляет собой один из локусов хромосомы. В нем образуется рРНК, которая затем образует комплекс с белками, формируя субъединицы рибосом, которые уходят в цитоплазму.
Негистоновые белки ядра образуют внутри ядра структурную сеть. Она представлена слоем фибрилл, подстилающим ядерную оболочку. К ней прикрепляется внутриядерная сеть фибрилл, к которой присоединены фибриллы хроматина.
Ядерная оболочка состоит из двух мембран: внешней и внутренней, разделенных межмембранным пространством. Внешняя мембрана соприкасается с цитоплазмой, на ней могут находиться полирибосомы, а сама она может переходить в мембраны эндоплазматического ретикулума. Внутренняя мембрана связана с хроматином. Таким образом, ядерная оболочка обеспечивает фиксацию хромосомного материала в трехмерном пространстве ядра.
Оболочка ядра имеет круглые отверстия – ядерные поры (рис. 7). В области поры внешняя и внутренняя мембраны смыкаются и образуют отверстия, заполненные фибриллами и гранулами. Внутри поры располагается сложная система из белков, обеспечивающих избирательное связывание и перенос макромолекул. Количество ядерных пор зависит от интенсивности метаболизма клетки.
Эндоплазматический ретикулум , или эндоплазматическая сеть (ЭПР), представляет собой причудливую сеть каналов, вакуолей, уплощенных мешков, соединенных между собой и отделенных от гиалоплазмы мембраной (рис. 8).
Различают шероховатый и гладкий ЭПР. На мембранах шероховатого ЭПР находятся рибосомы (рис. 9), которые синтезируют белки, экскретируемые из клетки или встраивающиеся в плазматическую мембрану. Вновь синтезированный белок сходит с рибосомы и проходит через специальный канал внутрь полости эндоплазматического ретикулума, где он подвергается посттрансляционной модификации, например связыванию с углеводами, протеолитическому отщеплению части полипептидной цепи, образованию S–S-связей между остатками цистеина в цепи. Далее эти белки транспортируются в комплекс Гольджи, где входят либо в состав лизосом, либо секреторных гранул. В обоих случаях эти белки оказываются внутри мембранного пузырька (везикулы).
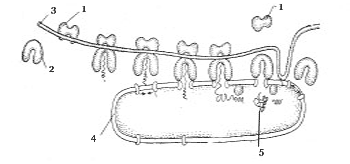
Рис. 9. Схема синтеза белка в
шероховатом ЭПР: 1 – малая и
2 – большая субъединицы рибосомы; 3 – молекула
рРНК;
4 – шероховатый ЭПР; 5 – вновь синтезируемый
белок
Гладкий ЭПР лишен рибосом. Его основная функция – синтез липидов и метаболизм углеводов. Он хорошо развит, например, в клетках коркового вещества надпочечников, где содержатся ферменты, обеспечивающие синтез стероидных гормонов. В гладком ЭПР в клетках печени находятся ферменты, осуществляющие окисление (детоксикацию) чужеродных для организма гидрофобных соединений, например лекарств.

Рис. 10. Аппарат Гольджи: 1 – пузырьки; 2 – цистерны
Комплекс Гольджи (рис. 10) состоит из 5–10 плоских ограниченных мембраной полостей, расположенных параллельно. Концевые части этих дискообразных структур имеют расширения. Таких образований в клетке может быть несколько. В зоне комплекса Гольджи находится большое количество мембранных пузырьков. Часть из них отшнуровывается от концевых частей основной структуры в виде секреторных гранул и лизосом. Часть мелких пузырьков (везикул), переносящих синтезированные в шероховатом ЭПР белки, перемещается к комплексу Гольджи и сливается с ним. Таким образом комплекс Гольджи участвует в накоплении и дальнейшей модификации продуктов, синтезированных в шероховатом ЭПР, и их сортировке.
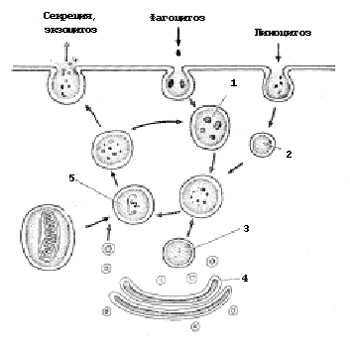
Рис. 11. Образование и функции лизосом: 1 – фагосома; 2 – пиноцитозный пузырек; 3 – первичная лизосома; 4 – аппарат Гольджи; 5 – вторичная лизосома
Лизосомы – это вакуоли (рис. 11), ограниченные одной мембраной, которые отпочковываются от комплекса Гольджи. Внутри лизосом достаточно кислая среда (рН 4,9–5,2). Там располагаются гидролитические ферменты, расщепляющие различные полимеры при кислых рН (протеазы, нуклеазы, глюкозидазы, фосфатазы, липазы). Эти первичные лизосомы сливаются с эндоцитозными вакуолями, содержащими компоненты, которые должны расщепляться. Вещества, попавшие во вторичную лизосому, расщепляются до мономеров и переносятся через мембрану лизосомы в гиалоплазму. Таким образом, лизосомы участвуют в процессах внутриклеточного переваривания.
Митохондрии окружены двумя мембранами: наружной, отделяющей митохондрию от гиалоплазмы, и внутренней, отграничивающей ее внутреннее содержимое. Между ними располагается межмембранное пространство шириной 10–20 нм. Внутренняя мембрана образует многочисленные выросты (кристы ). В этой мембране располагаются ферменты, обеспечивающие окисление образовавшихся за пределами митохондрий аминокислот, сахаров, глицерина и жирных кислот (цикл Кребса) и осуществляющие перенос электронов в дыхательной цепи (схема). За счет переноса электронов по дыхательной цепи с высокого на более низкий энергетический уровень часть освобождающейся свободной энергии запасается в виде АТФ – универсальной энергетической валюты клетки. Таким образом, основная функция митохондрий – это окисление различных субстратов и синтез молекул АТФ.
Схема переноса двух электронов по дыхательной цепи
Внутри митохондрии находится кольцевая молекула ДНК, которая кодирует часть белков митохондрии. Во внутреннем пространстве митохондрий (матриксе) находятся рибосомы, похожие на рибосомы прокариот, которые и обеспечивают синтез этих белков.
Тот факт, что митохондрии имеют свою кольцевую ДНК и прокариотические рибосомы, привел к возникновению гипотезы, согласно которой митохондрия является потомком древней прокариотической клетки, когда-то попавшей внутрь эукариотической и в процессе эволюции взявшей на себя отдельные функции.
Рис. 12. Хлоропласты (А) и тилакоидные мембраны (Б)
Пластиды – органоиды растительной клетки, которые содержат пигменты. В хлоропластах содержится хлорофилл и каротиноиды, в хромопластах – каротиноиды, в лейкопластах пигментов нет. Пластиды окружены двойной мембраной. Внутри них располагается система мембран, имеющая форму плоских пузырьков, называемых тилакоидами (рис. 12). Тилакоиды уложены в стопки, напоминающие стопки тарелок. Пигменты встроены в мембраны тилакоидов. Их основная функция – поглощение света, энергия которого с помощью ферментов, встроенных в мембрану тилакоида, преобразуется в градиент ионов Н + на мембране тилакоида. Как и митохондрии, пластиды имеют собственную кольцевую ДНК и рибосомы прокариотического типа. По-видимому, пластиды также являются прокариотическим организмом, живущим в симбиозе с клетками эукариот.
Рибосомы –это немембранные клеточные органоиды, встречающиеся как в клетках про-, так и эукариот. Рибосомы эукариот больше по размеру, чем прокариотические, их размер составляет 25х20х20 нм. Состоит рибосома из большой и малой субъединиц, прилегающих друг к другу. Между субъединицами в функционирующей рибосоме располагается нить иРНК.
Каждая субъединица рибосомы построена из рРНК, плотно упакованной и связанной с белками. Рибосомы могут располагаться в цитоплазме свободно или быть связанными с мембранами ЭПР. Свободные рибосомы могут быть единичными, но могут образовывать полисомы, когда на одной нити иРНК располагается последовательно несколько рибосом. Основная функция рибосом – синтез белка.
Цитоскелет – это опорно-двигательная система клетки, включающая белковые нитчатые (фибриллярные) образования, являющиеся каркасом клетки и выполняющие двигательную функцию. Структуры цитоскелета динамичны, они возникают и распадаются. Цитоскелет представлен тремя типами образований: промежуточными филаментами (нити диаметром 10 нм), микрофиламент ы (нити диаметром 5–7 нм) и микротрубочками . Промежуточные филаменты – неветвящиеся белковые структуры в виде нитей, часто расположенные пучками. Их белковый состав различен в разных тканях: в эпителии они состоят из кератина, в фибробластах – из виментина, в мышечных клетках – из десмина. Промежуточные филаменты выполнят опорно-каркасную функцию.
Микрофиламенты – это фибриллярные структуры, расположенные непосредственно под плазматической мембраной в виде пучков или слоев. Они хорошо видны в ложноножках амебы, в движущихся отростках фибробластов, в микроворсинках кишечного эпителия (рис. 13). Микрофиламенты построены из сократительных белков актина и миозина и являются внутриклеточным сократительным аппаратом.
Микротрубочки входят в состав как временных, так и постоянных структур клетки. К временным относится веретено деления, элементы цитоскелета клеток между делениями, а к постоянным – реснички, жгутики и центриоли клеточного центра. Микротрубочки – это прямые полые цилиндры с диаметром около 24 нм, их стенки образованы округлыми молекулами белка тубулина. Под электронными микроскопом видно, что сечение микротрубочки образовано 13 субъединицами, соединенными в кольцо. Микротрубочки присутствуют в гиалоплазме всех эукариотических клеток. Одна из функций микротрубочек – создание каркаса внутри клеток. Кроме того, по микротрубочкам, как по рельсам, перемещаются мелкие везикулы.
Клеточный центр состоит из двух центриолей, расположенных под прямым углом друг к другу и связанных с ними микротрубочек. Эти органеллы в делящихся клетках принимают участие в формировании веретена деления. Основой центриоли являются расположенные по окружности 9 триплетов микротрубочек, образующих полый цилиндр, шириной 0,2 мкм и длиной 0,3–0,5 мкм. При подготовке клеток к делению центриоли расходятся и удваиваются. Перед митозом центриоли участвуют в образовании микротрубочек веретена деления. Клетки высших растений не имеют центриолей, но у них есть аналогичный центр организации микротрубочек.
Все организмы, которые имеют клеточное строение, относятся к одной из групп (надцарств) – прокариотам (предъядерным) или эукариотам (ядерным).
К эукариотам относятся царства грибов, растений и животных. В переводе с греческого языка слово «эукариот» означает «владеющий ядром», т. е. все эукариоты имеют ядро. Эукариотические клетки в целом по строению сходны. Хотя есть и заметные различия между клетками организмов, которые принадлежат к различным царствам живой природы. Например, растительные клетки имеют различные пластиды и крупную центральную вакуоль , которая иногда отодвигает ядро к периферии. В клетках грибов стенка, как правило, состоит из хитина, а пластиды отсутствуют. В клетках животных нет ни пластид, ни плотной стенки, ни центральной вакуоли.
Кроме достаточно крупных рибосом у эукариот имеется и много других органоидов: ЭПС, митохондрии, клеточный центр, пластиды и т. д.
Клетки прокариот имеют относительно простое строение . В них нет организованного ядра, а единственная хромосома не отделена мембраной от остальной части клетки , а лежит непосредственно в цитоплазме. Тем не менее, в этой хромосоме записана вся наследственная информация клетки. К прокариотам относятся бактерии, цианобактерии и архебактерии.
Цитоплазма прокариот очень бедна по составу структур. В ней находятся многочисленные мелкие рибосомы. Функциональную роль хлоропластов и митохондрий выполняют специальные мембранные складки.
Сами клетки эукариот и прокариот сильно отличаются и по размеру . Эукариотическая клетка превышает клетку прокариот по объему в 1000 раз, а по диаметру в 10 раз. Диаметр эукариотической клетки составляет 0,01-0,1 мм, а прокариотической – 0,0005-0,01 мм.
Эукариоты и прокариоты отличаются и по генетическому аппарату . Так, генетический аппарат эукариотической клетки находится в ядре и защищен оболочкой. ДНК эукариот линейная, в соотношении 50/50 соединена с белками. У них образуется хромосома. В отличие от эукариот, ДНК у прокариот кольцевая, голая (с белками почти не соединена), лежит в особой области цитоплазмы – нуклеоиде и отделяется от остальной цитоплазмы при помощи мембраны.
Эукариотическая клетка делится при помощи митоза, мейоза или комбинации данных способов. Жизненный цикл эукариот состоит из двух ядерных фаз. Первая (гаплофаза) отличается одинарным набором хромосом. Во второй фазе (диплофаза) две гаплоидные клетки, сливаясь, образуют диплоидную клетку, в которой содержится двойной набор хромосом. Через несколько делений клетка снова становится гаплоидной.
Подобный жизненный цикл не характерен для прокариот. В основном прокариоты размножаются простым делением.
Эукариоты, в отличие от прокариот, могут переваривать твердые частицы путем их заключения в мембранный пузырек. Считается, что следствием данного процесса (фагоцитоза) у эукариот стало появление первых хищников.
Эукариоты отличаются от прокариот и наличием двигательных приспособлений . Эукариоты имеют жгутики, которые обладают сложным строением. Жгутики представляют собой тонкие клеточные выросты, которые окружены тремя слоями мембраны. Данные выросты содержат девять пар микротрубочек по периферии и две в центре. Жгутики имеют толщину до 0,1 мм и способны изгибаться. Также, помимо жгутиков, эукариоты имеют реснички. Реснички и жгутики идентичны по своей структуре и отличаются только размером. Длина ресничек достигает не более 0,01 мм.
Для некоторых прокариот также характерно наличие жгутиков, толщина которых очень мала и составляет около 20 нанометров в диаметре. Жгутики прокариот представляют собой пассивно вращающиеся полые белковые нити.
Считается, что первыми около 3,5 млрд лет назад появились прокариоты, которые через 2,4 млрд лет положили начало возникновению эукариотических клеток.
Итак:
- К эукариотам относятся грибы, растения и животные, а к прокариотам – бактерии, цианобактерии и архебактерии.
- У эукариот любого царства ядро есть. Именно в ядре находится генетический аппарат эукариотов, защищенный специальной оболочкой. У прокариот ядра нет.
- Клетки прокариот имеют простое строение, а единственная хромосома со всей наследственной информацией лежит просто в цитоплазме, в отличие от клетки эукариот, которая гораздо сложнее и многообразнее.
- Цитоплазма прокариот бедна по составу и имеет много мелких рибосом. У эукариот имеются крупные рибосомы и еще много других органоидов. Сама клетка по объему превышает клетку прокариот в 1000 раз, а по диаметрув 10 раз.
- ДНК эукариот линейная, наполовину соединена с белками и в ней образуется хромосома. У прокариот ДНК кольцевая, голая и лежит в нуклеоиде – особой области цитоплазмы.
- Эукариоты в основном размножаются посредством митоза и мейоза или сочетанием обоих способов, а прокариоты размножаются делением клетки надвое.
Важнейшая, основополагающая особенность эукариотических клеток связана с расположением генетического аппарата в клетке. Генетический аппарат всех эукариот находится в ядре и защищён ядерной оболочкой (по-гречески «эукариот» значит имеющий ядро). ДНК эукариот линейная (у прокариот ДНК кольцевая и находится в особой области клетки - нуклеоиде, который не отделён мембраной от остальной цитоплазмы). Она связана с белками-гистонами и другими белками хромосом, которых нет у бактерий.
В жизненном цикле эукариот обычно присутствуют две ядерные фазы (гаплофаза и диплофаза). Первая фаза характеризуется гаплоидным (одинарным) набором хромосом, далее, сливаясь, две гаплоидные клетки (или два ядра) образуют диплоидную клетку (ядро), содержащую двойной (диплоидный) набор хромосом. Иногда при следующем делении, а чаще спустя несколько делений клетка вновь становится гаплоидной. Такой жизненный цикл и в целом диплоидность для прокариот не характерны.
Третье, пожалуй, самое интересное отличие, - это наличие у эукариотических клеток особых органелл, имеющих свой генетический аппарат, размножающихся делением и окружённых мембраной. Эти органеллы - митохондрии и пластиды. По своему строению и жизнедеятельности они поразительно похожи на бактерий. Это обстоятельство натолкнуло современных учёных на мысль, что подобные организмы являются потомками бактерий, вступившими в симбиотические отношения с эукариотами. Прокариоты характеризуются малым количеством органелл, и ни одна из них не окружена двойной мембраной. В клетках прокариот нет эндоплазматического ретикулума, аппарата Гольджи, лизосом.
Ещё одно важное различие между прокариотами и эукариотами - наличие у эукариот эндоцитоза, в том числе у многих групп - фагоцитоза. Фагоцитозом (дословно «поедание клеткой») называют способность эукариотических клеток захватывать, заключая в мембранный пузырёк, и переваривать самые разные твёрдые частицы. Этот процесс обеспечивает в организме важную защитную функцию. Впервые он был открыт И. И. Мечниковым у морских звезд. Появление фагоцитоза у эукариот скорее всего связано со средними размерами (далее о размерных различиях написано подробнее). Размеры прокариотических клеток несоизмеримо меньше, и поэтому в процессе эволюционного развития эукариот у них возникла проблема снабжения организма большим количеством пищи. Как следствие среди эукариот появляются первые настоящие, подвижные хищники.
Большинство бактерий имеет клеточную стенку, отличную от эукариотической (далеко не все эукариоты имеют её). У прокариот это прочная структура, состоящая главным образом из муреина (у архей из псевдомуреина). Строение муреина таково, что каждая клетка окружена особым сетчатым мешком, являющимся одной огромной молекулой. Среди эукариот клеточную стенку имеют многие протисты, грибы и растения. У грибов она состоит из хитина и глюканов, у низших растений - из целлюлозы и гликопротеинов, диатомовые водоросли синтезируют клеточную стенку из кремниевых кислот, у высших растений она состоит из целлюлозы, гемицеллюлозы и пектина. Видимо, для более крупных эукариотических клеток стало невозможно создавать клеточную стенку из одной молекулы высокую по прочности. Это обстоятельство могло заставить эукариот использовать иной материал для клеточной стенки. Другое объяснение состоит в том, что общий предок эукариот в связи с переходом к хищничеству утратил клеточную стенку, а затем были утрачены и гены, отвечающие за синтез муреина. При возврате части эукариот к осмотрофному питанию клеточная стенка появилась вновь, но уже на другой биохимической основе.
Разнообразен и обмен веществ у бактерий. Вообще всего выделяют четыре типа питания, и среди бактерий встречаются все. Это фотоавтотрофные, фотогетеротрофные, хемоавтотрофные, хемогетеротрофные (фототрофные используют энергию солнечного света, хемотрофные используют химическую энергию).
Важнейшая, основополагающая особенность эукариотических клеток связана с расположением генетического аппарата в клетке. Генетический аппарат всех эукариот находится в ядре и защищён ядерной оболочкой. ДНК эукариот линейная (у прокариот ДНК кольцевая и находится в особой области клетки -нуклеоиде, который не отделён мембраной от остальной цитоплазмы). Она связана с белками-гистонами и другими белками хромосом, которых нет у бактерий.
В жизненном цикле эукариот обычно присутствуют две ядерные фазы (гаплофаза и диплофаза). Первая фаза характеризуется гаплоидным (одинарным) набором хромосом, далее, сливаясь, две гаплоидные клетки (или два ядра) образуют диплоидную клетку (ядро), содержащую двойной (диплоидный) набор хромосом. Иногда при следующем делении, а чаще спустя несколько делений клетка вновь становится гаплоидной. Такой жизненный цикл и в целом диплоидность для прокариот не характерны.
Третье отличие - это наличие у эукариотических клеток особых органелл, имеющих свой генетический аппарат, размножающихся делением и окружённых мембраной. Эти органеллы - митохондрии и пластиды. По своему строению и жизнедеятельности они поразительно похожи на бактерий.. Прокариоты характеризуются малым количеством органелл, и ни одна из них не окружена двойной мембраной.
Ещё одно важное различие между прокариотами и эукариотами - наличие у эукариот эндоцитоза, в том числе у многих групп - фагоцитоза. Фагоцитозом называют способность эукариотических клеток захватывать, заключая в мембранный пузырёк, и переваривать самые разные твёрдые частиц. Размеры прокариотических клеток несоизмеримо меньше, и поэтому в процессе эволюционного развития эукариот у них возникла проблема снабжения организма большим количеством пищи. Как следствие среди эукариот появляются первые настоящие, подвижные хищники.
Большинство бактерий имеет клеточную стенку, отличную от эукариотической. Строение муреина таково, что каждая клетка окружена особым сетчатым мешком, являющимся одной огромной молекулой. Среди эукариот клеточную стенку имеют многие протисты, грибы и растения. У грибов она состоит из хитина и глюканов, у низших растений - из целлюлозы и гликопротеинов, диатомовые водоросли синтезируют клеточную стенку из кремниевых кислот, у высших растений она состоит из целлюлозы, гемицеллюлозы и пектина. Видимо, для более крупных эукариотических клеток стало невозможно создавать клеточную стенку из одной молекулы высокую по прочности. Это обстоятельство могло заставить эукариот использовать иной материал для клеточной стенки. Другое объяснение состоит в том, что общий предок эукариот в связи с переходом к хищничеству утратил клеточную стенку, а затем были утрачены и гены, отвечающие за синтез муреина. При возврате части эукариот к осмотрофному питанию клеточная стенка появилась вновь, но уже на другой биохимической основе.
Разнообразен и обмен веществ у бактерий. Вообще всего выделяют четыре типа питания, и среди бактерий встречаются все. Это фотоавтотрофные, фотогетеротрофные, хемоавтотрофные, хемогетеротрофные. Эукариоты же либо сами синтезируют энергию из солнечного света, либо используют готовую энергию такого происхождения. Это может быть связано с появлением среди эукариотов хищников, необходимость синтезировать энергию для которых отпала.
Ещё одно отличие - строение жгутиков. У бактерий жгутиками являются полые нити диаметром 15–20 нм из белка флагеллина. Строение жгутиков эукариот гораздо сложнее. Они представляют собой вырост клетки, окруженный мембраной, и содержат цитоскелет (аксонему) из девяти пар периферических микротрубочек и двух микротрубочек в центре. В отличие от вращающихся прокариотическох жгутиков жгутики эукариот изгибаются или извиваются.
Две группы рассматриваемых нами организмов, как уже было сказано, сильно отличаются и по своим средним размерам. Диаметр прокариотической клетки составляет обычно 0,5–10 мкм, когда тот же показатель у эукариот составляет 10–100 мкм. Объём такой клетки в 1000–10000 раз больше, чем прокариотической.
Клетки эукариот содержат как более крупные рибосомы. Первые прокариоты возникли в процессе эволюции около 3,5 млрд. лет назад, от них около 1,2 млрд. лет назад произошли эукариотические организмы.
8.Бактерии - это одноклеточные организмы растительного происхождения, но лишенные хлорофилла. Они относятся к прокариотам, видны в световой микроскоп, размеры их измеряются в микрометрах. Бактерии растут на искусственных питательных средах, размножение происходит бинарным делением.
Бактерии делятся
вызывают болезни челове- вызывают заболевания не вызывают забо-
ка, животных, растений при определенных усло- леваний
Морфология - это форма, размер бактерий, расположение клеток в препарате. Различают три морфологических формы бактерий:
Различают три морфологических формы бактерий:
1) кокки 2) палочки 3) извитые
1. Кокки: форма - круглая
размер - мелкие
расположение в препаратах - 6 разновидностей:
а) микрококки б) диплококки в) тетракокки
гонококки пневмококки
г) сарцины в) стафилококки д) стрептококки
2. Палочковидные :
· форма - цилиндр
· размер: длина : толщина :
Крупные - толстые
Средних размеров - тонкие
· концы палочек – закругленные (кишечная палочка)
Прямые (сибиреязвенная палочка)
В виде утолщения (дифтерийная палочка)
· расположение - беспорядочное
В цепочку (стрептобактерии)
Попарно (диплобактерии)
В виде римских цифр II, V, X и т.д.
3. Извитые : форма – спиралевидная:
1. спириллы,
2. кампилобактерии
Бактериальная клетка состоит из клеточной стенки, цитоплазматической мембраны, цитоплазмы с включениями и ядерного аппарата, называемого нуклеоидом. Имеются другие структуры: мезосома, хроматофоры, тилакоиды, вакуоли, включения полисахаридов, жировые капельки, капсула (микрокапсула, слизь), жгутики, пили. Некоторые бактерии способны образовывать споры.
Структуру и морфологию бактерий изучают с помощью различных методов микроскопии: световой, фазово-контрастной, интерференционной, темнопольной, люминесцентной и электронной.
Клеточная стенка
В клеточной стенки грамположительных бактерий содержится небольшое количество полисахаридов, липидов, белков. Основным компонентом клеточной стенки этих бактерий является многослойный пептидогликан (муреин, мукопептид), составляющий 40-90% массы клеточной стенки. С пептидогликаном клеточной стенки грамположительных бактерий ковалентно связаны тейхоевые кислоты (от греч. teichos - стенка).
В состав клеточной стенки грамотрицательных бактерий входит наружная мембрана, связанная посредством липопротеина с подлежащим слоем пептидогликана. На ультратонких срезах бактерий наружная мембрана имеет вид волнообразной трехслойной структуры, сходной с внутренней мембраной, которую называют цитоплазматической. Основным компонентом этих мембран является бимолекулярный (двойной) слой липидов. Внутренний слой наружной мембраны представлен фосфолипидами, а в наружном слое расположен липополисахарид (ЛПС). Липополисахарид наружной мембраны состоит из трех фрагментов: липида А - консервативной структуры, практически одинаковой у грамотрицательных бактерий; ядра, или стержневой, коровой части (лат. core - ядро), относительно консервативной олигосахаридной структуры (наиболее постоянной частью ядра ЛПС является кетодезоксиоктоновая кислота); высоковариабельнои О-специфической цепи полисахарида, образованной повторяющимися идентичными олигосахаридными последовательностями (О-антиген). Белки матрикса наружной мембраны пронизывают ее таким образом, что молекулы белка, называемые поринами, окаймляют гидрофильные поры, через которые проходят вода и мелкие гидрофильные молекулы.
При нарушении синтеза клеточной стенки бактерий под влиянием лизоцима,
пенициллина, защитных факторов организма образуются клетки с измененной (часто шаровидной) формой: протопласты - бактерии, полностью лишенные клеточной стенки; сферопласты - бактерии с частично сохранившейся клеточной стенкой. Бактерии сферо- или протопластного типа, утратившие способность к синтезу пептидогликана под влиянием антибиотиков или других факторов и способные размножаться, называются L-формами.
Они представляют собой осмотически чувствительные, шаровидные, колбовидные клетки различной величины, в том числе и проходящие через бактериальные фильтры. Некоторые L-формы (нестабильные) при удалении фактора, приведшего к изменениям бактерий, могут реверсировать, «возвращаясь» в исходную бактериальную клетку.
Между наружной и цитоплазматической мембранами находится периплазматическое пространство, или периплазма, содержащая ферменты (протеазы, липазы, фосфатазы, нуклеазы, бета-лактомазы) и компоненты транспортных систем.
 Самый древний вид спорта в мире
Самый древний вид спорта в мире Владимир Колокольцев: служба в Афгане, борьба с ОПГ и другие факты биографии главы МВД
Владимир Колокольцев: служба в Афгане, борьба с ОПГ и другие факты биографии главы МВД Чем отличается лук шалот от репчатого лука, можно ли заменять их друг другом отличий от репчатого лука
Чем отличается лук шалот от репчатого лука, можно ли заменять их друг другом отличий от репчатого лука Все бабы как бабы, а я – дура на миллион И главный вопрос: какой женщиной для этого нужно быть
Все бабы как бабы, а я – дура на миллион И главный вопрос: какой женщиной для этого нужно быть Эльза скарлетт титания. Перевооружение эльзы. Сердечные тайны Эрзы Скарлетт
Эльза скарлетт титания. Перевооружение эльзы. Сердечные тайны Эрзы Скарлетт Персонаж аниме Эльза Алая: биография, доспехи и интересные факты
Персонаж аниме Эльза Алая: биография, доспехи и интересные факты Самая популярная аниме героиня
Самая популярная аниме героиня